张 政,卢鸿健,李明慧,高文博,田 宇,王嘉祺,张荣花,章广玲
(1.华北理工大学临床医学院/河北省医工融合精准医疗重点实验室,河北 唐山 063000;
2.华北理工大学基础医学院/河北省慢性疾病基础医学重点实验室,河北 唐山 063210)
肝细胞癌(hepatocellular carcinoma,HCC)是世界上常见的恶性肿瘤之一,占原发性肝癌的75%~85%。GLOBOCAN 的最新统计数据显示[1],2020 年全球HCC 新发病例达91 万,死亡病例83 万,分别居肿瘤新发病率第6 位、死亡率第3 位;
我国人口占世界的18%,但新发病例占全球的45%。HCC 的发生发展过程复杂,是一种异质性较高的癌症,大多数发现时已是中晚期,错过了手术的最佳时间且预后较差,其5 年总生存率只有12%[2]。因此,寻找HCC 发展过程中有效的靶点和信号通路并进行相应干预,对延缓HCC 的进展或者逆转具有重要意义。研究表明[3],非编码RNA(non-coding RNA,ncRNA)包括长链非编码RNA(long non-coding RNA,lncRNA)和微小RNA(microRNA,miRNA)等,其可能在HCC 的发生发展中发挥了重要的作用。miRNAs 分子由18~25个bp 组成的非编码RNA,能够通过碱基互补配对与靶基因的3" 端非编码区(3"untranslated region,3"UTR)特异性结合,沉默或降解靶mRNAs 从而影响其翻译功能。lncRNAs 分子是一种长度超过200 bp 的非编码核苷酸,含有miRNAs 的结合位点,可以作为miRNAs 的“海绵”吸附miRNAs,从而竞争性抑制miRNAs 对靶mRNAs 的作用[4]。本研究拟通过生物信息学方法探究癌症基因组图谱(TCGA)数据库中HCC 组织中差异表达的基因,并构建lncRNAmiRNA-mRNA 调控网络,为靶向诊断和治疗HCC提供科学依据。
1.1 HCC 组织中差异表达基因的筛选 利用R 软件DESeq2 包分析TCGA 数据库中374 例HCC 组织与50 例正常肝组织中lncRNAs、miRNAs、mRNAs 的表达谱。使用R 软件ggplot2 包对表达谱数据绘制火山图,以|Log2FC|>1(fold change 差异倍数大于2 倍)且P<0.05 作为筛选标准。
1.2 miRNAs 转录因子的预测 利用FunRich 软件预测差异表达miRNAs 的转录因子,根据其所调控的基因数和值进行排序,选取富集程度最显著的前10 个转录因子进行展示。
1.3 基因功能(GO)和京都基因与基因组百科全书(KEGG)富集分析 利用R 软件clusterProfiler 包分别分析miRNAs、mRNAs 各基因之间潜在的影响,并以调整后的P<0.05 为标准,进行GO 和KEGG分析,分别预测其主要生物功能和参与的主要信号通路。
1.4 miRNA-mRNA 调控网络 使用FunRich 软件预测miRNA 的候选靶基因,结合HCC 组织中差异表达的miRNAs 和mRNAs 的FC 值,筛选具有相互作用的miRNA-mRNA 关系。
1.5 PPI 网络构建 将筛选出的基因导入STRING 数据库,分析蛋白之间的相互作用(protein-protein interaction,PPI),并将分析结果导入Cytoscape 中,构建可视化PPI 蛋白互作网络图。
1.6 lncRNA-miRNA-mRNA 调控网络构建 利用Starbase 数据库,预测miRNAs 的上游lncRNAs,结合HCC 中差异表达lncRNAs 的FC 值,筛选具有相互作用的lncRNA-miRNA 关系对。基于共享的miRNAs 通过Cytoscape 软件对lncRNA-miRNAmRNA 调控网络进行可视化。
2.1 差异表达lncRNAs、miRNAs、mRNAs 的筛选 利用R 软件DESeq2 包分析374 例HCC 组织和50 例正常肝 组织中lncRNAs、miRNAs、mRNAs 的表达谱,根据P<0.05 和|Log2FC|>1 的标准筛选筛选出HCC 组织中差异表达的lncRNAs 2850 个,其中上调2475 个,下调375 个;
差异表达的miRNAs 38个,其中上调31 个,下调7 个;
差异表达的mRNAs 4455 个,其中上调3199 个,下调1256 个,见图1。

图1 差异表达基因的火山图
2.2 转录因子-miRNA 调控网络 利用FunRich 软件对38 个差异表达miRNAs 进行转录因子预测,共得到201 个转录因子,根据其所调控的基因数和P值,选取富集程度显著的前10 个转录因子进行展示,见图2,其基本信息见表1,前10 个转录因子分别为EGR1、SP1、NKX6-1、POU2F1、LHX3、SP4、MEF2A、ARID3A、MSX2、HOXA9。

表1 差异表达前10 名转录因子的基本信息

图2 差异表达miRNAs 转录因子的预测
2.3 miRNAs 的GO 和KEGG 富集分析 对38 个差异表达的miRNAs 进行GO 和KEGG 功能富集分析,其生物过程主要涉及血管内皮细胞迁移的调节、血管内皮细胞增殖调控与新生血管生成、内皮细胞增殖的调节、血管内皮细胞增殖与新生血管生成;
分子功能主要涉及mRNA 结合参与转录后基因沉默、RNA 聚合酶Ⅱ复合物结合、RNA 聚合酶核心酶结合、基础转录机制结合;
KEGG 分析主要富集于miRNAs 在癌症中的作用,见图3。

图3 差异表达miRNAs 的GO 和KEGG 功能富集分析
2.4 miRNA-mRNA 调控网络 通过FunRich 软件预测差异表达的38 个miRNAs 的下游靶基因,共得到1530 个靶基因,然后与差异表达的4455 个mRNAs取交集获得286 个候选靶基因,见图4。通过Starbase 和MiRDB 数据库比对分析,获得14 302 对miRNA-mRNA,与差异表达的miRNA-mRNA 匹配后,最后筛选出交叉表达负调控的12 个miRNAs 和108 个mRNAs 作为后续研究对象。
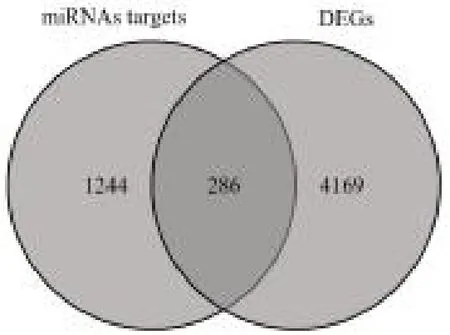
图4 差异表达miRNAs 的下游靶基因与差异表达mRNAs 的韦恩图
2.5 108 个mRNAs 的蛋白PPI 调控网络及富集分析利用STRING 数据库和Cytoscape 软件构建筛选出的108 个基因的PPI 网络,见图5。图中节点代表mRNAs,有相互作用关系的mRNAs 之间有连线,共有44 个节点和134 条边;
节点越大、颜色越深则介数中心度越大,边的粗细代表连接评分,边越粗,mRNAs 间的互作关系越大。

图5 108 个mRNAs 的蛋白互作网络图
2.6 108 个mRNAs 的GO 与KEGG 富集分析 利用GO 功能分析筛选出的108 个mRNAs 的生物功能,生物过程主要涉及肌肉适应、MAP 激酶活性的调节、胚胎器官发育、蛋白丝氨酸/苏氨酸激酶活性的正调控;
分子功能主要涉及MAP 激酶酪氨酸/丝氨酸/苏氨酸磷酸酶活性、MAP 激酶磷酸酶活性、激酶调节活性、DNA 结合转录激活物活性和RNA 聚合酶Ⅱ特异性。KEGG 富集分析108 个mRNAs 参与的信号通路,主要涉及调节干细胞多能性的信号通路、催乳素信号通路、MAPK 信号通路、EGFR 酪氨酸激酶抑制剂耐药性,见图6。
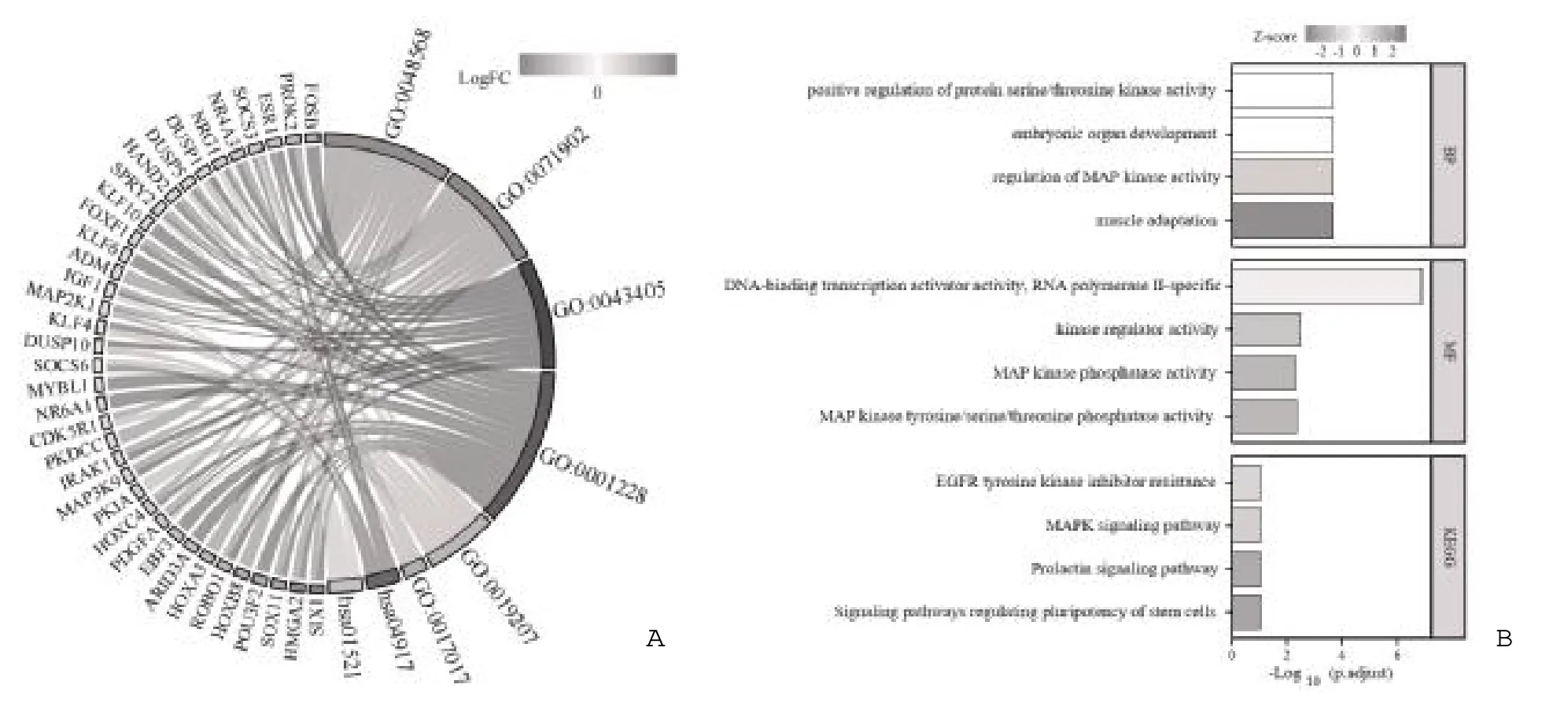
图6 108 个mRNAs 的GO 和KEGG 富集分析图
2.7 构建lncRNA-miRNA-mRNA 调控网络 利用Starbase 数据库,分析筛选出的12 个miRNAs 的上游lncRNAs 共221 个,结合差异表达lncRNAs 的FC 值,筛选出5 对lncRNA-miRNA,包括2 个miRNAs(miR-23A 和miR-146B)与5 个lncRNAs(GAS5、MCM3AP-AS1、LINC00173、KCNQ1OT1、SNHG7);
结合筛选出的2 个miRNAs 的与组织中差异表达mRNAs 的FC 值,共匹配42 对miRNAmRNA(包括40 个mRNAs)。通过Cytoscape 软件对lncRNA-miRNA-mRNA 调控网络进行可视化处理,见图7。

图7 HCC 组织中lncRNA-miRNA-mRNA 的调控网络
HCC 是由肝细胞恶性病变引起的癌症,患者发现时大多数已是中晚期,具有发病隐匿、进展快、易耐药、预后差以及易复发等特点,且现有的治疗效果并不理想[5]。因此寻找开发有效的HCC 诊断和预后的生物标记物具有重要意义。越来越多的研究表明,lncRNAs 和miRNAs 等非编码RNAs 与HCC 的发病密切相关。LncRNA n339260 通过降低miR-30e-5p的表达使TP53INP1 的表达增加,从而促进肝细胞癌的进展[6]。LncRNA ZEB1-AS1 通过调节miR-1224-5p/MAP4K4 轴,调节持续沙眼衣原体感染中线粒体介导的HeLa 细胞凋亡[7]。抑制lncRNA GAS6-AS2 通过调节miR-136-5p/OXSR1 轴能够在体外和体内水平减轻败血症相关的急性肾损伤[8]。然而在HCC 中尚没有报道完整的lncRNA-miRNA-mRNA 调控网络,因此深入研究lncRNAs 和miRNAs 在HCC 发生发展中的作用及调控机制具有重要意义。
已有研究表明,转录因子调控miRNAs 的表达,且这些转录因子在HCC 的发病过程中具有重要的调控作用。本研究预测了调控HCC 中差异表达miRNAs 的上游转录因子共201 个,根据值筛选出差异显著的前10 个转录因子分别为EGR1、SP1、NKX6-1、POU2F1、LHX3、SP4、MEF2A、ARID3A、MSX2、HOXA9,其中EGR1 差异最明显。EGR1 是一种转录因子,可能参与肿瘤细胞的增殖、侵袭和转移以及肿瘤血管生成,与癌症的发生和发展密切相关[9]。有报道表明[10],EGR1 可能以转录因子方式调控hsa-miR-146b 的表达。EGR1 在miR-195 的上游启动子区有结合位点,且调控miR-195 的表达,EGR1 可抑制miR-195/AKT3 轴并抑制胃癌细胞凋亡[11]。EGR1 还能够通过负调控miR-491-5p 促进细胞增殖、生长和迁移,并抑制细胞凋亡,从而抑制胃癌的进程[12]。因此,研究转录因子对miRNAs 的调控作用可在系统层次上对疾病的发生提供理论依据。
近年来,伴随着高通量测序技术的发展产生了大量的基因数据,这为进一步研究HCC 的发生发展及预后监测提供了有力的基础。本研究首先分析TCGA 数据库中差异表达基因的表达谱,得到的差异表达的lncRNAs 2850 个、miRNAs 38 个、mRNAs 4455 个。对38 个差异表达的miRNAs 进行GO 和KEGG 功能富集分析,显示主要涉及血管内皮细胞迁移和增殖、新生血管生成、内皮细胞增殖的调节等生物过程,mRNA 结合参与转录后基因沉默等分子功能,且KEGG 分析显示这些miRNAs 主要在癌症中发挥作用。通过FunRich 软件预测得到的miRNAs 靶基因与差异表达的mRNAs 交叉匹配,对筛选出的108 个mRNAs 进行GO 分析发现,主要在胚胎器官发育、MAP 激酶活性的调节等生物学过程富集,DNA 结合转录激活物活性和RNA 聚合酶Ⅱ特异性、激酶调节活性等分子功能富集。KEGG 分析发现,靶基因参与的信号通路主要涉及调节干细胞多能性的信号通路、MAPK 信号通路等。因此可以得出在HCC 中差异表达的mRNAs 其生物功能主要与细胞增殖、形态分化和细胞迁移有关。同样地,在PPI 网络的Top 10 基因中KLF4、NRG1、FOSB、MAP2K1 也参与了上述生物学功能以及相关信号通路。研究发现[13],miR-146B 通过靶向KLF4 调节肝星状细胞活化。NRG1 是鳞状细胞癌分化的关键调节因子,抑制肿瘤细胞角化和终末鳞状分化并促进肿瘤细胞增殖[14]。TGF-β1通过诱导FOSB 促进前列腺癌细胞迁移和侵袭[15]。CircRNA ZFR 通过上调MAP2K1 促进HCC 的增殖[16]。
本研究寻找与miRNAs 相匹配的上游lncRNAs和下游mRNAs 构建HCC 中lncRNA-miRNA-mRNA 的调控网络,其中包含2 个关键的miRNAs(miR-23A、miR-146B)以及与其匹配的5 个lncRNAs(GAS5、MCM3AP-AS1、LINC00173、KCNQ1OT1、SNHG7)和40 个mRNAs(ROBO1、LIN28B 等)。研究表明[17],miR-23A 是HCC 中下调最显著的miRNA,可能是改善肝癌患者索拉非尼反应性的潜在靶点。miR-146B 通过靶定PTP1B 抑制胃癌细胞的增殖并促进其凋亡[18]。miR-146B 通过NF-κB(核因子κB)途径靶向TLR4(Toll 样受体4),抑制胆囊癌肿瘤的发生和转移[19]。以上研究表明,miR-23A、miR-146B具有抑制肿瘤的作用。此外,lncRNA GAS5 靶向miR-23A 抑制CCl4 诱导的肝纤维化[20]。LncRNA SNHG7 通过调节miR-146B-5p/ROBO1 轴促进胰腺癌进展[21]。LncRNA SNHG17 通过抑制miR-23A-3p 来调节CXCL12 介导的血管生成,从而促进结直肠腺癌细胞的增殖和迁移[22]。LncRNA KCNQ1OT1通过miR-146A-5p/ACER3 轴抑制HCC 的放射敏感性并促进肿瘤发生[23]。
综上所述,lncRNA-miRNA-mRNA 调控网络的构建,有望阐明HCC 的发生发展机制,为HCC 的临床诊断和预后监测提供有价值的标记物。但是lncRNA-miRNA-mRNA 调控网络的功能以及各差异基因的功能,还有待通过体内外实验和临床实验进一步证实。
猜你喜欢激酶调控调节方便调节的课桌青少年科技博览(中学版)(2022年11期)2023-01-07蚓激酶对UUO大鼠肾组织NOX4、FAK、Src的影响天津医科大学学报(2021年3期)2021-07-21蚓激酶的药理作用研究进展世界科学技术-中医药现代化(2021年12期)2021-04-192016年奔驰E260L主驾驶座椅不能调节汽车维修与保养(2021年8期)2021-02-16如何调控困意学苑创造·A版(2020年12期)2020-01-07经济稳中有进 调控托而不举中国外汇(2019年15期)2019-10-14顺势而导 灵活调控作文教学研究(2016年1期)2016-07-05可调节、可替换的takumi钢笔工业设计(2016年4期)2016-05-04黏着斑激酶和踝蛋白在黏着斑合成代谢中的作用中国医药生物技术(2015年4期)2015-12-26SUMO修饰在细胞凋亡中的调控作用医学研究杂志(2015年8期)2015-06-22